细胞染色质结构变化调控遗传信息的开放程度,从而影响基因活性,改变细胞命运和细胞状态。这一过程在发育、免疫应答、肿瘤发生发展等方面具有重要作用。利用ATP的能量,染色质重塑复合物滑动、弹出、交换或解聚核小体,实现对染色质结构的动态调控。其中,ISWI家族染色质重塑复合物ISW1a不仅能滑动核小体,还能感知周围染色质环境,促进酵母细胞形成等间距排布的核小体阵列。ISW1a主要聚集在基因组的启动子区,通过滑动核小体来关闭启动子染色质。
2024年1月5日,清华大学生命科学学院/北京生物结构前沿研究中心/清华-北大生命科学联合中心陈柱成教授研究团队在《自然结构与分子生物学》(Nat. Struct. Mol. Biol)杂志在线发表题为“ISW1a复合物结合双核小体的结构”(Structure of the ISW1a complex bound to the dinucleosome)的研究论文。该论文首次报道了染色质重塑复合物结合双核小体的结构,揭示了ISW1a催化核小体等间距排布的分子机制,为理解染色质重塑在高级染色质环境中发挥功能提供了新的认识角度。
ISW1a复合物由马达亚基Isw1和调控亚基Ioc3组成,图a。通过ISW1a结合底物核小体的结构可以看到,ISW1a的两个亚基分别识别一个核小体,图b。其中Isw1马达亚基结合的核小体可以发生滑动,称为N1核小体。Isw1亚基与N1核小体的结合机制有三处,分别是马达结构域结合核小体的DNA和H4尾巴,“精氨酸锚”(arginine anchor, RA )结合核小体酸性表面(acidic patch, AP),以及HSS结构域结合入口端接头DNA。Ioc3亚基结合的核小体称为N2核小体,结合位点也有三处,分别是入口端接头DNA、核小体的H4尾巴,以及核小体酸性表面。HSS-Ioc3共同形成一个DNA结合模块,识别入口端长度~30 bp的接头DNA。马达结构域和HSS-Ioc3模块协同作用于N1核小体,揭示了ISW1a作为“蛋白标尺”,推动间距为30 bp的核小体阵列的分子机制。此外,该研究首次发现核小体阵列中邻近的核小体参与调控染色质重塑活性。Ico3结合N2核小体,促进了ISW1a的核小体间距活性,图c。这种对邻近核小体的识别作用在DNA损伤修复(图d),以及基因组启动子区核小体的规则排布(图e)等方面扮演重要角色。
结合课题组之前关于RSC染色质重塑复合物的研究结果(Ye,Science 2019),我们可以发现,尽管ISW1a与RSC都识别启动子DNA,但由于两者三维空间结构的不同,使得染色质重塑马达结构域分别结合+1位核小体的两个不同表面。这种马达定位的差异导致+1位核小体滑向两个相反方向,从而发挥出相互制衡的功能, 图f。具体来说,ISW1a将+1位核小体滑向上游启动子区,抑制基因的转录;而RSC把+1位核小体滑向下游,打开启动子区,促进基因的转录。因此,这两大类染色质重塑复合物一开和一关,共同调控基因表达的精细平衡。
清华大学生命科学学院陈柱成教授为本文通讯作者,清华大学生命科学学院2018级博士生李莉菲(已毕业)和2022级博士后陈康净、2020级博士生谢悠扬为该论文共同第一作者,生命学院2019级博士生胡鹏晶、郑州大学叶佑丕研究员也参与了重要工作。本工作获得国家自然科学基金、科技部重大科学研究计划专项、北京市结构生物学高精尖创新中心、清华-北大生命科学联合中心、国家蛋白质科学研究(北京)设施清华基地的大力支持,国家蛋白质科学研究(北京)设施清华基地冷冻电镜平台和计算平台为数据收集和处理提供了支持。
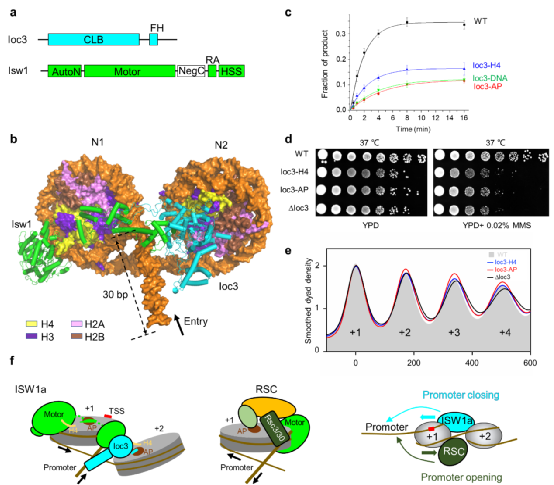
图1 ISW1a复合物结合双核小体的结构和相关功能研究
(a)ISW1a复合物的亚基组成。 (b)ISW1a结合双核小体的电镜结构。黑箭头表示DNA移动的方向。 (c) Ioc3野生型与突变体复合物的核小体间距排布活性。 (d)野生型与Ioc3突变体酵母菌株的生长表型分析。(e)野生型与Ioc3突变体酵母菌株基因的启动子附件+1~+4位核小体阵列排布。(f)ISW1a 与RSC滑动启动子核小体,相互制衡的工作模型。TSS, 转录起始位点;AP, 酸性表面。青色和深绿色弯箭头分别表示ISW1a 与RSC对启动子DNA的结合,直箭头表示对应的+1核小体的移动方向。
原文连接:
https://www.nature.com/articles/s41594-023-01174-6
Structure of the RSC complex bound to the nucleosome | Science